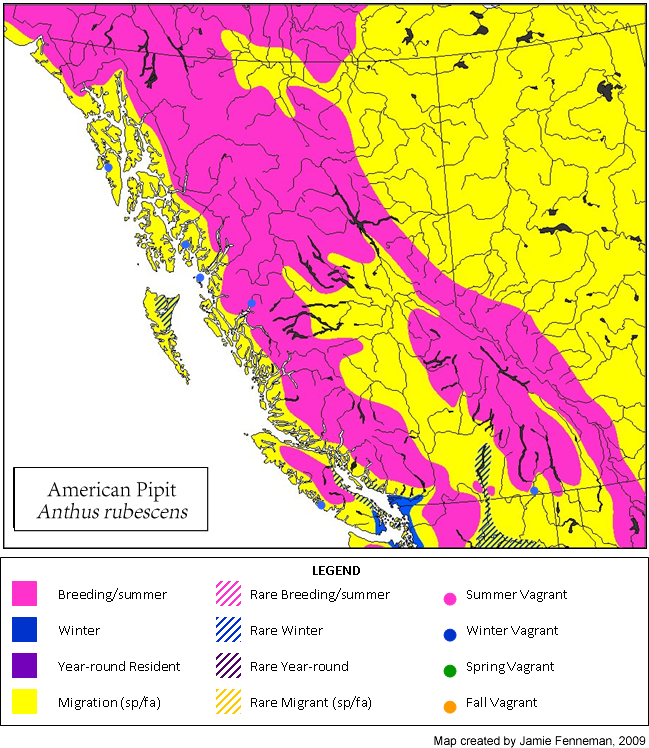
The American Pipit is likely to be confused only with other pipits, all of which are vagrants to British Columbia. Non-breeding and juvenile Red-throated Pipit, which is a rare migrant along the coast, is distinguished from American Pipit by its pale-streaked upperparts (giving the impression of pale ‘braces’ on the mantle), more prominent whitish wing bars, much bolder and more extensive blackish streaking on the underparts, heavier blackish malar stripe, and pink legs. Breeding-plumaged adults, which have variable (though usually extensive) pinkish-red to red on the head and upper breast are very distinctive and are unlikely to be confused with American Pipit. Sprague’s Pipit, which is a prairie species that is casual in the interior of B.C. (but has bred), is overall much paler and buffier, with a dark crown and distinctly unmarked and plain facial pattern that highlights the prominent dark eye. In addition, the upperparts are heavily streaked with black and buff and the underparts are almost unstreaked. Sprague’s Pipit is also shorter-tailed than American Pipit, giving it a noticeably different structure.
| The male’s song, which is usually (though not always) given in flight, is a slow series of high, repeated, clear or jingling phrases, often with different phrases given during the ascending phase of the display flight than are given during the descending phase; phrases are also usually given more rapidly during the descending phase. Song transcriptions for include chueet.…..chueet…..chueet……chueet……chueet……chueet and churee….. churee…..churee…..churee…..churee. Territorial males also give a rapid piptitititititi and a slower twet-twet-twet-tea-tea-tea-twet-twet during territorial disputes on the breeding grounds. Agitated individuals give a low, rising pwisp when a predator approaches the nest. The most commonly-heard calls throughout the year, and the only calls heard outside of the breeding season, are a high-pitched, chirping, single- or double-noted slip or slip-ip (commonly sounding like pip-it), often interspersed with a higher, lisping tseeep or tsitsiip. The call notes of A.r.japonicus are slightly higher-pitched than those of the North American subspecies, and the call is usually given as a single (rather than doubled) note. Source: Verbeek and Hendricks (1994); Sibley (2000); Rosenberg (2006a) |
Courtship
Pairs usually form on the breeding grounds, although some individuals may pair up during spring migration. Males begin performing territorial song flights shortly after arrival on the breeding grounds. During these displays, which usually last 10-30 seconds, the male gradually climbs (often steeply) into the air while singing, remains for several seconds in the air while continuing to sing, and then descends to the ground with the wings held open and the tail feathers spread while singing a more rapid series of phrases than that which was given during the ascending phase of the display. Song flights can attain heights of up to 40 m above the ground, although there is considerable variation in the height of singing birds. The male also sings from the ground prior to performing the song flight.
Nest
Nest building commences immediately after pair formation. Both sexes contribute to the building of the nest, which takes an average of 5 days to complete. The nest is placed on the ground, usually partially protected by overhanging vegetation, rocks, or soil. It is a small cup (6-8 cm across, 3-5 cm deep) of dry grasses, sedges, leaves, moss, rootlets, lichens, and plant stems that is placed in a shallow hollow in the ground and is lined with fine grasses and hair.
Eggs
A single clutch of (2) 4-6 (7) eggs is laid in June (rarely late May) or early July and is incubated by the female for 14-15 days before hatching. Eggs are present in B.C. between late May and late July. The smooth, glossy eggs are whitish-grey to buffy-white and are sparsely to heavily spotted with brown and pale grey, sometimes with thin black streaks or with accumulated dark markings around the larger end of the egg forming a ‘wreath’. This species is not known to be a host for Brown-headed Cowbird parasitism.
Young
The young are fully altricial upon hatching, with pinkish skin and a sparse covering of long brownish-grey to blue-grey down (shorter and whiter on the underside); the mouth is reddish-orange and the gape flanges are pale yellow. Both parents tend to the nestlings (although only the female broods), which remain in the nest for 14-16 days before fledging. The fledglings are tended by the parents for an additional 2 weeks after leaving the nest, after which time they become partially independent for another 4 weeks before dispersing and becoming fully independent. Nestlings and dependent fledglings are present in B.C. between mid-June and mid-August.
Source: Verbeek and Hendricks (1994); Baicich and Harrison (1997); Campbell et al. (1997)
|
This species forages almost exclusively on the ground, where it is commonly observed walking or running (rather than hopping) while constantly bobbing its tail. It consumes terrestrial and aquatic invertebrates (insects, caterpillars, spiders, marine worms, crustaceans, etc.) and seeds which it picks from the surface of the soil or (in the case of many invertebrates) gleans from low vegetation. In aquatic habitats, it sometimes wades into shallow water to pick at organisms on the surface of the water. During winter and migration, it is commonly attracted to habitats such as sewage ponds, piles of rotting seaweed, or the edges of alkaline wetlands where there is a high abundance insects (especially flies). It forages alone during the breeding season, but during winter and migration it is usually encountered in small to very large, loose flocks that often number into the hundreds of individuals.
Source: Verbeek and Hendricks (1994)
|
|
.jpg){kind=link}